17 顔
ヒトの「顔」の起源を探る
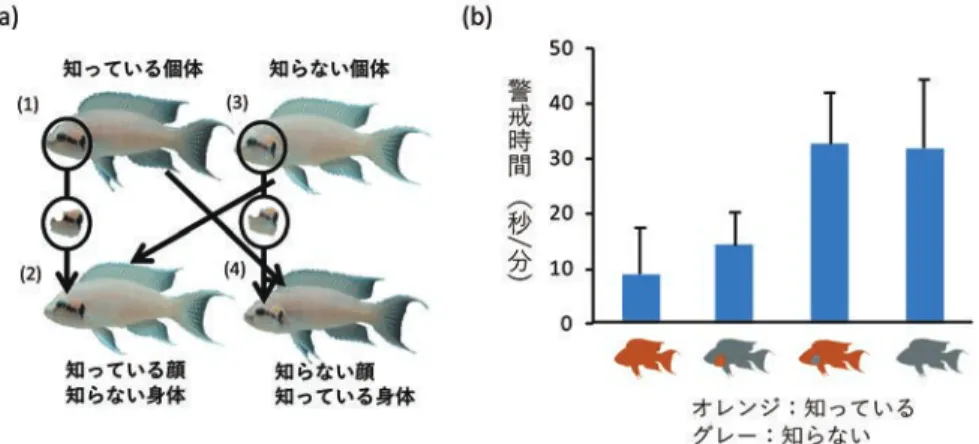
私たちヒトにとって「顔」は特別な視覚刺激 であると考えられている(Leopold & Rhodes, 2010)。「顔」は他の身体部位と比較して多様 な情報を含んでおり,性別や年齢だけではな く感情や人種などを知ることができる。さら に,顔の個人間の違いはわずかであるにもか かわらず,素早く正確に個人を識別すること ができる。また,顔の情報を処理するときに パーツごとの違いに注目するのではなく,顔全 体として違いを検出する「全体処理(holistic processing)」 と 呼 ば れ る 顔 特 異 的 な 処 理 を していることが報告されている(Leopold & Rhodes, 2010)。このような高度な処理を必要 とするヒトの顔認知(face recognition)はど のように進化してきたのだろうか。顔認知の進 化について明らかにするためには,ヒト以外の 動物が「顔」をどのように知覚し,どのような 情報を獲得しているのかというアプローチが有 効である。本稿ではヒトと系統的には離れた分 類群であるサカナの「顔」に関する認知能力に 焦点を当てた研究について紹介する。 サカナの「顔」に含まれる情報 水族館で優雅に泳ぐサカナたちの顔を 注意深く観察したことはあるだろうか。 よくよく見れば目の近くには模様があっ たり,個体ごとに異なる顔を持っていた りすることがわかるだろう。特にサンゴ 礁に生息するスズメダイやベラの仲間で は色鮮やかな模様がある種が多い(Kohda et al., 2015)。そこでまずは私たちが顔の特徴で他 人を識別できるように,サカナたちが眼の近く にある模様で他個体を識別しているのかについ て調べた。 アフリカにはタンガニイカ湖という九州 ほどの大きさの湖があり,シクリッドと呼 ばれるカワスズメ科に属するサカナたちが 生活している。このシクリッドの一種である Neolamplorogus pulcherというサカナ(図1a) は眼の近くにオレンジや茶色,黒色や青色で構 成された複雑な模様をもち,その模様は個体 ごとに異なることがわかっている。そこでこ のサカナの写真を撮影し,「知っている個体」, 「知っている個体の顔+知らない個体の身体」, 「知らない個体」,「知らない個体の顔+知って いる個体の身体」という四つのモデルを作成 し(図2a),それらのモデルに対する反応を調 べた。すると,「身体」が知っている個体のも のかどうかにかかわらず,「知らない個体の顔」 に対する警戒時間が長いことがわかった(図 2b; Kohda et al., 2015)。この結果はN. pulcher
ヒトの「顔」とサカナの「顔」
日本学術振興会 特別研究員PD堀田 崇
(ほった たかし) Profile─ 大阪市立大学大学院理学研究科生物地球系専攻で学位(理学)取得。日本学術振興 会特別研究員 PD として,大阪市立大学動物機能生態学研究室を経て,現在は京都大学心理学研究室に在籍。専門 は動物行動学,動物心理学。論文は「魚類を対象とした比較認知科学研究の可能性」(動物心理学研究),The use of multiple sources of social information in contest behavior(共著,Frontiers in Ecology and Evolution)など。図 1 顔で他個体を識別していることが明らかにされたサカ ナ。(a)Neolamprologus pulcher(Kohda et al., 2015) (b)ディ スカス(Symphysopdon aequifasciatus;Satoh et al., 2016) (c)Julidochromis transcriptus(Hotta et al., 2018)。
18 et al., 2017)。このように,まるでヒトの表情 のように顔色を変化させることで刻々と変わる 内部状態を伝えているのである。 サカナは「顔」をどのように知覚する? 近年のサカナの「顔」に関する研究は,顔 からどのような情報を得ているのかについて 調べたものが多い。一方で,サカナたちが顔 をどのように知覚しているのかという知見 も,「顔認知」を理解するうえでは重要であ る。Kawasakaら(2019)は,サカナたちが他 個体の「顔」をヒトと同様に全体処理してい るのか,それともパーツごとに部分処理してい るのかを明らかにするために,倒立効果(face inversion effect)について調べた。倒立効果と は,顔が上下逆さまにして呈示されるとその認 識や識別が難しくなるという現象である。倒立 効果が生じるということは,顔を知覚するとき に個々のパーツとしてではなく顔全体として 認識していることの証拠であると考えられてお り,霊長類やヒツジなどの哺乳類で報告されて いる(Leopold & Rhodes, 2010)。そこで先ほ ど紹介したN. pulcherに同種2個体の頭部の写 真を正立像または倒立像として呈示した。その 結果,倒立像で呈示されたときは2個体の写真 を識別することができないことがわかった。こ のような現象は他の物体に対しては観察され なかったことから,N. pulcherがヒトと同様に が眼の近くにある模様で他個体を識別してい ることを示す。さらにこの識別が0.5秒以内に できるということも明らかになった。同様の結 果は熱帯魚であるディスカス(Symphysopdon aequifasciatus;図1b)やタンガニイカ湖に住む 他のシクリッド(Julidochromis transcriptus; 図1c)でも報告されている(Satoh et al., 2016; Hotta et al, 2018)。これらの種はN. pulcherと は異なり,全身に模様があるにも関わらず顔の 模様で個体識別をしているのである。また,一 見して顔に目立った模様の見られないメダカ でも顔(身体の前側の部分)で個体を識別で きることがわかっている(Wang & Takeuchi, 2017)。 サカナたちの「顔」には個体を識別するため の情報だけではなく,他にもいろいろな情報を 伝えていることが明らかとなっている。サン ゴ礁に棲むネッタイスズメダイ(Pomacentrus amboinensis)とニセネッタイスズメダイ(P. moluccensis)はヒトから見ると外見がとても よく似ている。しかし紫外線を当てると「顔」 に 複 雑 な 模 様 が 浮 か び 上 が る。Siebeckら (2010)は,実験の結果,この模様が種の認識 に用いられていることを明らかにした。さらに サカナたちは,模様によって種を認識している だけではなく,色の濃淡を変えることで闘争に 対するモチベーションや自身の順位をも他個体 に伝えていることも報告されている(Balzarini
図 2 N. pulcherを対象とした実験の概要(Kohda et al., 2015)。知っている個体と知らない個体の顔と身体 の組み合わせから 4 種類のモデル(a)を作成し,実験個体に呈示した。その結果,身体が知っているかど うかにかかわらず,知っている顔に対する警戒時間は知らない個体に対する警戒時間よりも短かった(b)。 Kohda et al.(2015)を一部改変。
19 顔 顔特異的な知覚をしていることが示唆された (Kawasaka et al., 2019)。「顔」を個々のパー ツではなく,全体として処理をすることは,よ り瞬時に多様な情報を獲得することを可能にし ているだろう。また顔を他の物体から検出しや すくしているとも考えられる。実際ヒトは三つ の点があれば一見してその模様が顔のように見 えてしまい,そこに注意が向いてしまう(シ ミュラクラ現象, Leopold & Rhodes 2010)。 ヒトやチンパンジーでは,様々な動物(哺乳 類)の写真を呈示したときに,まず顔を注視 することがわかっている(Kano & Tomonaga, 2009)。このような顔に対する選択的な注意は サカナにもみられるのだろうか。そこで私は, N. pulcherに同種他個体と全身写真を呈示し, どこの部分(顔・胴体・尾)をはじめに注視す るのかを調べた。すると彼らはまず顔に対し て注意を向けることがわかった(Hotta et al., 2019)。しかしN. pulcherは眼の近くの部位に しか目立った模様がない(図1a)ため,模様 の顕著性によって注意を向けているだけかも しれないという可能性が残る。そこで眼の周 辺のみならず全身にストライプ模様があるJ. transcriptusの全身写真(図1c)を提示したと ころ,同種のときと同様に顔に注意を向けるこ とがわかった。このような効果は楕円などの物 理的図形に起こらなかった。これらの結果か ら,ヒトやチンパンジーと同様にN. pulcherは まず顔に注意を向けるということが明らかに なった(Hotta et al., 2019)。 サカナの「顔」から顔認知の起源を探る これまで脊椎動物における顔認知の進化 についてはあまり議論されてこなかったが, LeopoldとRhodes(2010)ではヒトから無脊椎 動物までの顔認知研究についてレビューした 論文で1つの仮説を提唱している。その仮説と は,脊椎動物が誕生したころに眼や口が捕食者 や種の認識にとって重要な刺激であり,その後 それらを含む部位を「顔」として認識しはじめ たというものである。実際,海水魚であるデバ スズメダイ(Chromis caeruleus)は捕食者か どうかを「眼の大きさや口の形」で判断してい ることが明らかとなっている(Karplus et al., 1982)。しかしこれまで紹介してきた研究は, サカナたちもヒトや類人猿と同様に「顔」を検 出,知覚し,様々な情報を獲得していることを 示唆しているため,その進化について再考する 必要があるだろう。 近年の研究から哺乳類のみならずサカナも顔 から個体情報をはじめとする様々な情報を得て いるということが明らかになったが,それでは そのような顔認知の進化についてはどのように 考えればよいだろうか。言い換えれば,サカ ナと哺乳類の共通祖先に同等の顔認知能力が あったと言えるのだろうか,ということであ る。この問いに対してはっきりと答えをだすこ とは現状得られている限られた知見だけではむ ずかしいが,より詳細に哺乳類とサカナの顔を 比べてみるとその糸口は見つけられるかもしれ ない。特に個体識別に関する情報について考 えてみると,実は哺乳類とサカナの間には相 違点がある。哺乳類はパーツごとのわずかな違 いやパーツ間の距離や配置をもとに個体を識別 している(Leopold & Rhodes, 2010)が,サカ ナたちは「模様」を使って個体を識別している (Kohda et al., 2015; Satoh et al., 2016; Hotta et
al., 2018)。 もし「模様」を使うのであればそれが「顔」 である必要はないように思われる。Satohら (2016)は,サカナたちの個体識別の情報とし て顔にある模様が使われるようになった理由に ついて3つの仮説を提唱している。一つ目の仮 説は,サカナどうしが出会ったときに一般的に 頭部が向かい合うhead-to-headの姿勢をとるか らであるというものである("Encountering"仮 説)。しかしディスカスは正面顔ではなく横顔 でのみ他個体を識別しているということから この仮説は支持されないだろう(Satoh et al., 2016)。二つ目の仮説は,重要な社会的シグナ ルはヒレなど損傷しやすい部位ではなく身体の 中心部にあるというものである("Main-body" 仮説)。ただ必ずしもそれが顔である必要はな く,また全身に模様のある種でも「顔」で個体 ヒトの「顔」とサカナの「顔」
20
識別のシグナルとなっているということから, この仮説も考えにくい(Hotta et al., 2018)。む しろ身体の模様は性淘汰によるもの,もしく は隠蔽のためのものであると考えられている (Satoh et al., 2016; Hotta et al., 2018)。最後の 仮説は,サカナにとって「まず顔(特に眼)を 注視する」傾向があり,その副産物として眼の 周囲に重要な社会シグナルが集まったというも のである(“Face-specific(gazing-eye)”仮説)。 先述の通り,N. pulcherは他個体の写真を呈示 されたときにまず顔を注視することがわかっ ている(Hotta et al., 2019)。つまりLeopoldと Rhodes(2010)で提唱されたように,脊椎動 物が誕生したときには,顔(特に眼や口などの パーツ)はすでに種認識や捕食者検知のために 重要なシグナルとして用いられていた。そのた めに他個体と出会ったときにまずは顔を注視す るようになり,重要な社会シグナルが顔に多く 含まれるようになったという仮説である。サカ ナの顔認知研究はまだまだはじまったばかりで 十分な知見がないため,これ以上これらの仮説 について考察することはできない。しかし,全 身に模様のない種での検討や,サカナにおける 顔を呈示した時にどのような神経回路や脳部位 が反応しているのかを調べることにより顔認知 の進化について検証することができるだろう。 おわりに 本稿ではサカナの顔認知に関する研究を紹介 した。一見無表情に見えるサカナの顔にも多く の情報が含まれており,それらの複雑な情報を 巧みに読み取っていることがわかってきた。と きには私たちのように表情を変えてコミュニ ケーションをとっているのかもしれない。サ カナは地球上の様々な環境に適応しており,そ の社会性も多様であるということから,脊椎動 物における顔認知の進化にどのような社会要因 (e.g., 群れで生活するような高い社会性)が影 響しているのかについて検証するにはとてもい い分類群であろう。サカナの顔認知研究は近年 注目されはじめたばかりであり,今後どのよう な成果が報告されていくのかとても楽しみであ る。ぜひ水族館に足を運んだときには,優雅に 泳ぐサカナたちの顔をゆっくりと眺めてみては いかがだろうか。 文 献
Balzarini, V., et al. (2017). Computer animations of color markings reveal the function of visual threat signals in Neolamprologus pulcher. Current Zoology, 63, 45-54.
Hotta, T., et al. (2018). Face recognition in the Tanganyikan cichlid Julidochromis transcriptus. Animal Behavior, 127, 1-5.
Hotta, T., et al. (2019). Fish focus primarily on the faces of other fish. Scientific Reports, 9, 8377.
Kano, F. & Tomonaga, M. (2009). How chimpanzees look at pictures: A comparative eye-tracking study. Proceedings of the Royal Society B: Biological Sciences, 276, 1949-1955.
Karplus, I., et al. (1982). A preliminary experimental analysis of predator face recognition by Chromis caeruleus (Pisces, Pomacentridae). Zeitschrift fur Tierpsychologie 58, 53-65.
Kawasaka, K., et al. (2019). Does a cichlid fish process face holistically? Evidence of the face inversion effect. Animal Cognition, 22, 153-162.
Kohda, M., et al. (2015). Facial recognition in a group-living cichlid fish. PLoS ONE, 10, e0142552.
Leopold, D. A. & Rhodes, G. (2010). A comparative view of face perception. Journal of Comparative Psychology, 124, 233-251.
Satoh, S., et al. (2016). Facial recognition in a discus fish (Cichlidae): Experimental approach using digital models. PLoS ONE, 11, e0154543.
Siebeck, et al. (2010). A species of reef fish that uses ultraviolet patterns for covert face recognition. Current Biology, 20, 407-410.
Wang, M. Y. & Takeuchi, H. (2017). Individual recognition and the 'face inversion effect' in medaka fish (Oryzias latipes). eLife, 6, e24728.